About This Item
- Full TextFull Text(subscription required)
- Pay-Per-View PurchasePay-Per-View
Purchase Options Explain
Share This Item
The AAPG/Datapages Combined Publications Database
AAPG Special Volumes
Abstract
Pub. Id:
First Page:
Last Page:
Book Title:
Article/Chapter:
Subject Group:
Spec. Pub. Type:
Pub. Year:
Author(s):
Abstract:
Observations off Bonaire, Netherlands Antilles, support the recent contention that Caribbean reefs are occasionally subjected to low-water emergence and are therefore more similar to reefs of the Indo-Pacific than previously believed.
Off Bonaire, some incipient reef flats and the shallow floor between the flats and shore are sporadically awash for extended periods of time. Most of the corals (representatives of over 20% of all known Caribbean species) that inhabit the emergent biotopes appear to be able to endure subaerial exposure without severely deleterious effects. However, destruction of Montastrea annularis either results directly from exposure or, more probably, is initiated by it.
Bonaire's shallow-water reefs are of three types. The type that occurs in turbulent water is composed almost entirely of Acropora palmata and is therefore similar to the typical, shallow-water reefs of the Caribbean. The two types that are found in calm water differ considerably in species composition from typical Caribbean reefs and differ from one another as a consequence of whether they undergo or escape emergence. Calm-water reefs that are constantly immersed consist almost exclusively of nearly continuous colonies of Montastrea annularis. Those that are intermittently emergent are the result of a succession of heterogeneous corals. Construction of the reefs of varied corals was begun by abundant large colonies of M. annularis, but most of these colonies were killed when they attained sizes at which their summits were subaerially exposed during exceptionally low stands of the sea. Following the decline of the M. annularis colonies and erosion of their coralla by boring organisms, construction of the reef framework was continued by an extraordinarily diverse coral assemblage. Framework building has been culminated upward by the establishment of exposure-tolerant corals and other cnidarians, but the diverse assemblage is maintained on the reef flanks and deeper portions of the flats.
The diversity of the corals (32 of the 48 species found off Bonaire) that are involved in the formation of the sometimes emergent reefs is unusual in shallow water in the Caribbean; such diversity is usually associated only with much deeper reef zones. Because Montastrea annularis has been shown to be dominant over most of the corals of the unusually diverse, shallow-water reef assemblage in the extracoelenteric-digestion hierarchy (i.e., a "pecking order" in which representatives of subordinate coral species are consumed and thereby eliminated from competition for reef space by those that are more aggressive), the diversity of the assemblage is attributed to the initial decimation and continued control by emergence of the once prolific colonies of this coral.
Text:
INTRODUCTION
Although exposure of a reef off the Tortugas during an unusually low tide was recorded by Vaughan (1919) in a pioneering synthesis of knowledge of reefs and reef corals, until recently, subsequent observations supported the generally held belief (Wells, 1957) that Caribbean reefs, unlike those of the Indo-Pacific, do not ordinarily undergo emergence. In the past few years, however, several reports have called attention to the rather common occurrence of low-water emergence of some Caribbean reefs and to the ability of the various coral species found on the reefs to survive exposure. In contrast, most accounts of emergence that were published in the years between Vaughan's survey and these recent reports have been limited to observations of occasional exposure of portions of reefs that do not include coral-inhabited biotopes or of coral assemblages that consist of only a single species. Moreover, many of the accounts that have noted exposure of individual species have not included observations of the consequences of the exposure.
Numerous investigators have recorded the growth of Siderastrea radians in seemingly inhospitable habitats, including ones from which water is occasionally withdrawn, but the careful study of the species by Yonge (1935) most clearly substantiates the
End_Page 155------------------------
ability of these corals to withstand brief intervals of exposure. Investigation of the tolerance to exposure of another species, Acropora palmata, apparently has not been undertaken, although casual reference has been made to the fact that the tops of colonies are sometimes awash (e.g., Newell et al, 1951, 1959; Newell and Rigby, 1957; Roos, 1964; Storr, 1964). Incidental mention only (Kissling, 1965) has also been made of emergence off the Florida Keys of a nonreef (nonrigid frame) assemblage consisting almost entirely of Porites furcata, but others have recorded observations that attest to the tolerance of this species to exposure. Porter (1974) has observed that off the Atlantic coast of Panama P. furcata survived a 1-hour interval of exposure at midday w thout apparent damage. Glynn (1973b) found that during a 7-year period off the southwestern coast of Puerto Rico, no large-scale kills of P. furcata occurred from repeated episodes of exposure, some of which took place at midday. Glynn has, however, implicated inordinately low tides--that occurred prior to his surveillance of the P. furcata assemblages--in the widespread mortality evidenced by clusters of dead corals in some areas.
Even before observations had been made of the ability of a few Caribbean corals to survive natural emergence, investigations had been carried out by Vaughan (1913) to determine the relative hardiness of a number of species that were experimentally exposed to the air. Vaughan found that nearly all of the species that he removed from shallow water off the Tortugas were able to withstand such treatment for an hour in the shade, and several survived for 4 hours. As might be expected, the endurance of the same corals in the sun was less. Vaughan's experimentation with artificially exposed corals demonstrated that Porites porites, P. astreoides, and Favia fragum, followed closely by Siderastrea radians and Manicina areolata, have the greatest cap city to withstand emergence. Therefore, it is not surprising that these corals, with the exception of M. areolata (which is not a frame builder in shallow water), are among those that have been mentioned as occupants of emergent reef zones in the recent reports describing exposure of assemblages of varied corals.
Excluding Vaughan's (1919) record of the emergent Tortugas reef, widespread exposure of several Caribbean reef-coral zones was first noted on atolls located off Nicaragua by Milliman (1969). Subsequently, Glynn (1973a) called attention to emergence of various coral species living on a flat formed by fossil sections of fringing reefs along the Caribbean coast of Panama, and I have previously mentioned exposure of Bonaire's reefs (Scatterday, 1974).
In summaries of current knowledge of Caribbean reefs, both Glynn (1973a) and Milliman (1973) have suggested that reef emergence is rather commonplace in at least some parts of the Caribbean, and the latter has contended that in the southern Caribbean, where they would never be subjected to the trauma of occasionally cool air temperatures, many different corals are able to survive low-water exposure. Although Glynn (1973a) has cited cases that substantiate the somewhat common occurrence of Caribbean reef emergence, in a number of these cases, corals are not present in the exposed reef zones or, if present, they do not comprise varied reef-building assemblages. Moreover, as the foregoing review of pertinent literature demonstrates, observations of exposure of heterogeneous assemblages h ve been recorded in only three recent reports. One of these (Scatterday, 1974) contains only brief mention of exposure of such assemblages and was unknown to Milliman (1973) at the time his summary was prepared. Therefore, further documentation of Milliman's contention seems desirable. For this reason, detailed descriptions are presented herein of the emergent reef zones off Bonaire, one of the three islands that comprise the Leeward group of the Netherlands Antilles (see paper by Bak, this volume). In addition, the suggestion is made in a subsequent section that emergence is an important factor in determining the unusually diverse composition of one of Bonaire's reef-coral assemblages.
NATURE OF EMERGENCE OFF BONAIRE
With the exception of more recently discovered reefs made up of laterally continuous colonies of living Montastrea annularis, Bonaire's reefs and associated coral assemblages
End_Page 156------------------------
have been fully described elsewhere (Scatterday, 1974); hence, only brief descriptions are presented here.
Fringing reefs are found off Bonaire's entire leeward coast, but shallow reefs that reach heights sufficient to allow partial emergence at times of extreme low water are discontinuous, and the widely separated stretches along which they occur would, if joined end-to-end, extend only half the length of the coast. Although such shallow reefs have been constructed by two distinctly different assemblages of corals, the upward termination of their growth at the same elevation has led to the formation of morphologically similar incipient reef flats. In some areas, the flats, like those common along the windward sides of many Caribbean islands and banks, are formed by the accordant level reached by the tops of tall colonies of Acropora palmata. As might be expected from the usual wi dward position of such flats, they are restricted to parts of Bonaire's leeward coast where wave activity is strong enough to promote the exceedingly rapid rate of growth that enables A. palmata to preempt all available space. In calm water, an assemblage consisting of an unusually large number of different coral species has been responsible for reef construction, and the reef flats are the result of upbuilding to a level surface through the colonization of successively higher dead-coral substrates. Construction was begun by Montastrea annularis, as evidenced by the fact that truncated coralla of now mostly dead, but once extremely large and presumably healthy colonies of this species lie at the base of most portions of the reefs. The relatively few remaining colonies o living M. annularis are small in size. Most result from continued growth of a few lobes located on the periphery of large, otherwise dead, colonies; however, some are secondary colonies that developed on dead portions of predecessors after settling. By far the greatest part of the extensive foundation area provided by the truncated M. annularis coralla is covered by an extraordinary variety of other corals. Many of those with fragile coralla have been broken and in turn covered by new colonies of the same or different species. Exceedingly diverse corals inhabit "pools" within the flats or occupy the reef flanks that descend from the flats in all directions (i.e., not only landward and seaward, but also toward the channels that transect the reefs at intervals along the oast). On the flats, however, live only the exposure-tolerant corals listed in Table 1, but zoanthiniarians and the hydrozoan Millepora may also be present. Colonies of Millepora are especially abundant on the seaward side of the reef flats where they may form a crest similar to the "Millepora zone" observed by Milliman (1969) on atolls off Nicaragua or the "Millepora sill" found by Glynn (1968, 1973b) on the seaward edge of reefs off Puerto Rico.
Seaward of the shallow reefs or, where the reefs are absent, beginning at depths between 3 and 4 m, are found the deeper-water coral zones that have been recognized in most reefs of the Caribbean region (i.e., in order of increasing depth, the Acropora cervicornis Zone, the Montastrea annularis Zone, a zone of abundant Montastrea cavernosa and/or Solenastrea bournoni, and a zone dominated by agariciids). Landward of the shallow reefs is a zone (equivalent to the "rear" and "shore" zones in the commonly used terminology of Goreau, 1959) of abundant Diploria clivosa that has also been generally recognized throughout the region. The Diploria clivosa Zone is equally as shallow (0 to 1 m) as the reef flats; hence, during times of low wa er, corals of both biotopes are exposed.
Commonly, during emergence (i.e., at occasional low spring tides and/or when exceptional winds drive or draw the water from the coast), coral colonies are sporadically immersed by intermittently higher waves, but at times they protrude for long intervals, up to 2/3 m above troughs or even crests of waves of average amplitude. While exposed, however, some portions of the colonies may be bathed by splash, to which Milliman (1969) has attributed survival of corals on the emergent reefs he has described. Numerous individual corals, which were first observed being subjected to these emergent conditions in late March 1971, have been examined over a 3-year period. Throughout this time, no deleterious effects of exposure on originally healthy colonies have been observed. Hiatuses in skeletal rowth may have occurred during
End_Page 157------------------------
Click to view image in JPEG format. Fig. 1. [Grey Scale] Emergence of corals in the Diploria clivosa Zone off Bonaire at noon on March 27, 1974. Clearly visible are numerous colonies of Acropora palmata. Small, hemispherical objects nearer shore are colonies of Diploria clivosa and D. strigosa. During 5-day period, March 27-31, low water was accompanied by calm conditions, and as result corals were exposed without being externally wetted for several midday hours each day. View to north; outskirts of Kralendijk, Bonaire's capital, can be seen beyond partially exposed reef flat in left background.
{kind=link}
Table 1. Living Caribbean Reef Corals Observed During Emergence
End_Page 158------------------------
exposure, but evidence of growth increments over emergent periods has not yet been studied.
Somewhat less commonly, low-water emergence coincides with exceedingly calm seas, and as a consequence the upper 60 cm of tall colonies or entire small ones are not externally wetted in any manner for several hours at a time (Figs. 1-5). Such exposure occurred for between 2 and 3 midday hours each day over a 5-day period in late March 1974. Throughout emergence, corals remained moist and glistening from exuded mucus, by which they presumably escaped severe desiccation. No morphologic changes indicative of damage were observed. Unfortunately, observations could be continued for only 1 week following return to normal-water conditions, but during this time none of the colonies that had been randomly selected while emergent for subsequent study showed any noticeable change. However, episo es of prolonged, midday exposure accompanied by negligible water agitation must at times have been responsible for the death of polyps occupying the tops of larger, massive coralla, especially those of Diploria strigosa (Fig. 3). Because of their smaller size or growth in a slightly deeper location, other colonies of D. strigosa have been somewhat less frequently exposed or have been awash for shorter intervals and therefore bear healthy polyps over their entire surfaces. Continued peripheral growth of truncated colonies has resulted in "microatolls" (Fig. 4) identical to those commonly formed by Porites lutea on reef flats in the Indo-Pacific. As there is scant detritus in the Diploria clivosa Zone or on the reef flats where the "microatolls" are loca ed, they are clearly not the result of sedimentation on the tops of once hemispherical colonies, as are the "microatolls" that have been previously recorded (Zaneveld, 1957) in deeper water off the Netherlands Antilles.
In spite of some evidence off Bonaire of damage to corals during past intervals of inordinately severe, low-water conditions, it is evident that a number of Caribbean corals can withstand several hours of exposure, even without external wetting, equally as successfully as their counterparts on the emergent portions of Indo-Pacific reefs. However, it must be emphasized that, unlike the rather regular tidal exposure of some
Click to view image in JPEG format. Fig. 2. [Grey Scale] Close view of several emergent colonies of Acropora palmata exhibiting form commonly called "panshoal" that is typical of specimens inhabiting Diploria clivosa Zone. Same time, date, and locality as Figure 1. Coralla are up to several meters across.
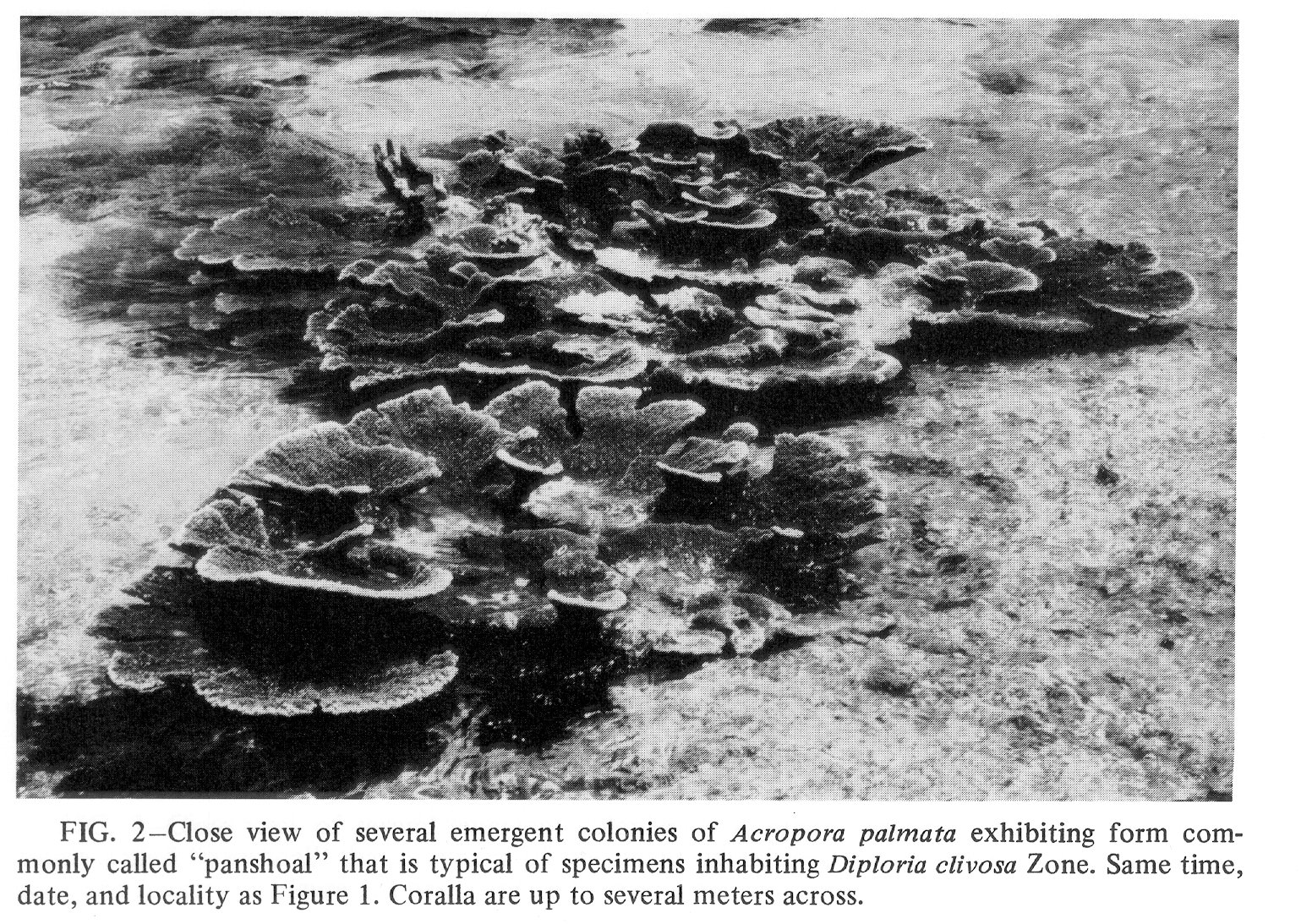{kind=link}
End_Page 159------------------------
Click to view image in JPEG format. Fig. 3. [Grey Scale] Close view of partially emergent colony of Diploria strigosa. Uppermost polyps have been killed, presumably during earlier episode of unusually severe exposure, and bioerosion has leveled summit of the corallum. Same time, date, and locality as Figure 1. Corallum diameter is approximately 0.5 m.
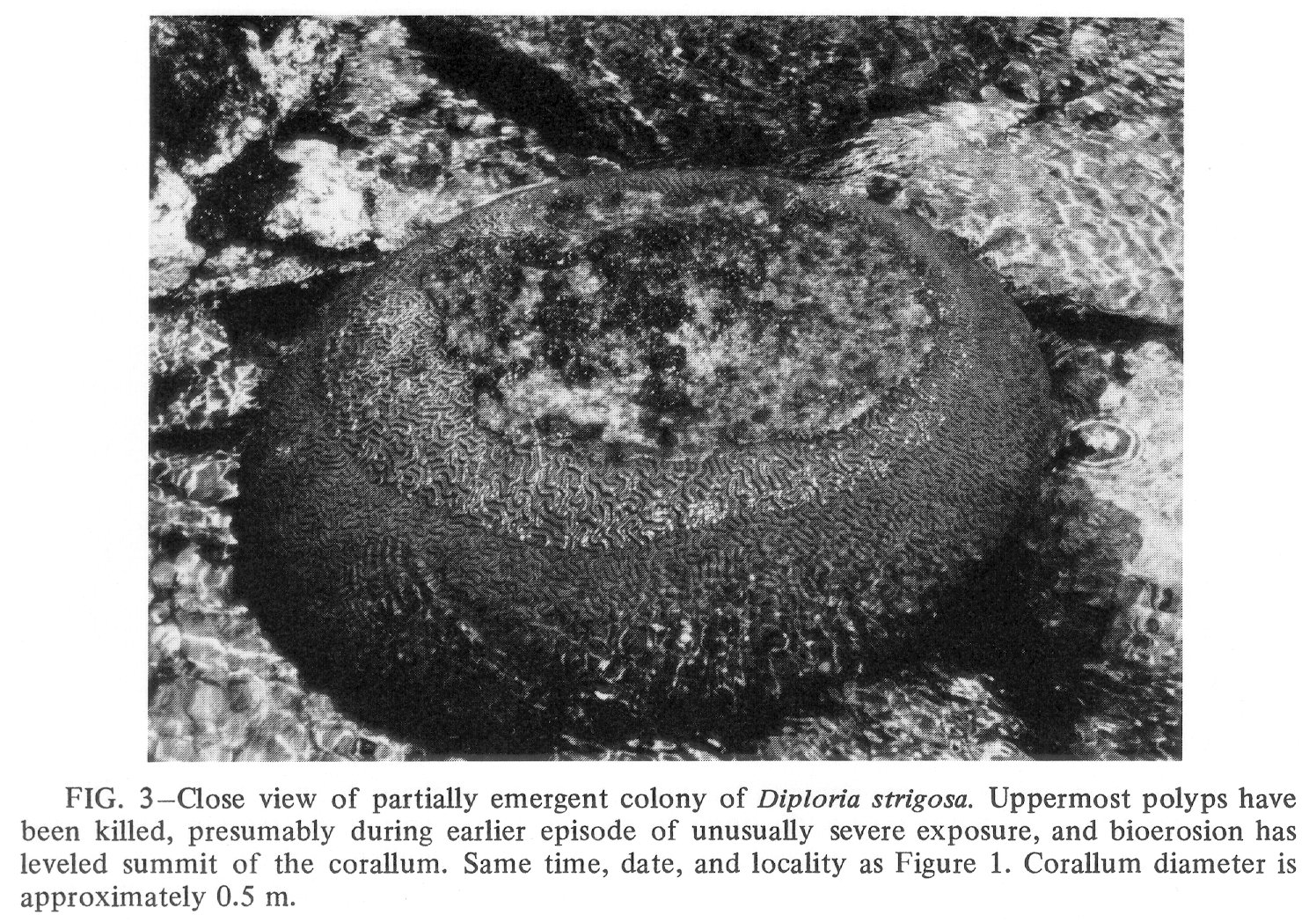{kind=link}
Click to view image in JPEG format. Fig. 4. [Grey Scale] Seaward part of incipient reef flat that is beginning to be slowly inundated following several hours of exposure. "Microatoll" that appears near center has developed as result of continued peripheral growth of truncated colony of Diploria strigosa like that illustrated in Figure 3. Although colonies of Millepora complanata occur scattered over the flat, they are concentrated near seaward edge where they form a "Millepora sill," a portion of which appears beyond the "microatoll." Same date and locality as Figure 1; time approximately 2:30 p.m. "Microatoll" diameter is approximately 2 m.
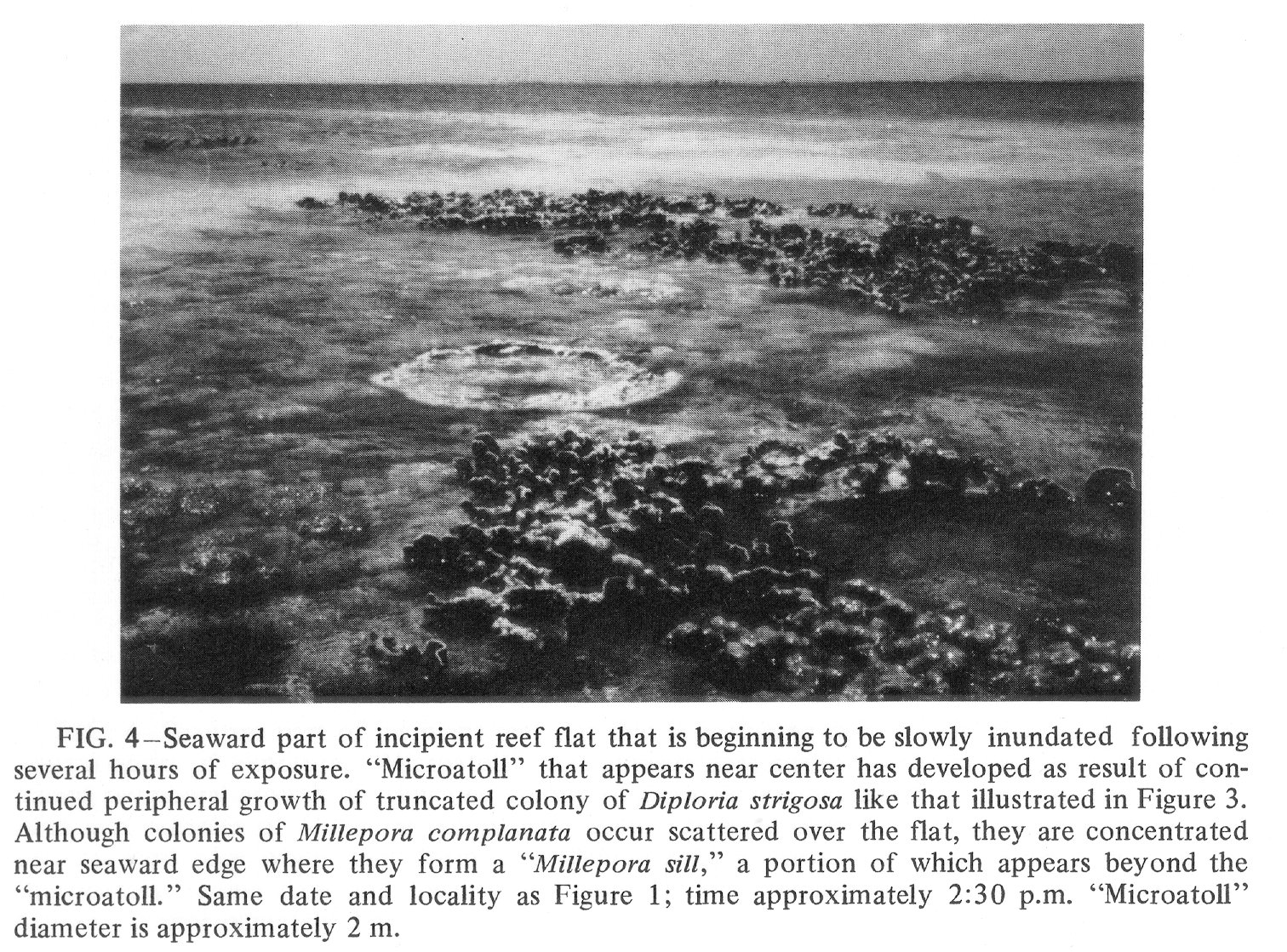{kind=link}
End_Page 160------------------------
Indo-Pacific corals, corals off Bonaire are subjected to infrequent, sporadic exposure separated by many months of total immersion.
The timing of low-water emergence off Bonaire is more irregular and thus less predictable than might be anticipated from the general regime of extreme low tides that Glynn (1968) has established from tide-table data for the other two islands of the Netherlands Antilles Leeward group. The modifying effects of local meteorologic conditions on the tidal regime off Bonaire appear to be more comparable to those that Glynn (1972) has observed off the Pacific coast of Panama.
EFFECTS OF EMERGENCE ON MONTASTREA ANNULARIS
Excluding from count the 12 ahermatypes that number among the 66 scleractinian corals now recognized in the Caribbean (Wells and Lang, 1973; Wells, 1973), slightly over 20% of all reef-building corals have been recorded in sometimes emergent biotopes, and another hermatype--Porites furcata--may be a significant constituent of the nonrigid-frame communities occupying reef flats that are occasionally awash. As indicated by Table 1, all of the reef-building corals that have been observed undergoing emergence in other areas have been seen in this situation off Bonaire, with the notable exception of Montastrea annularis. This coral is completely absent from the Diploria clivosa Zone off Bonaire, and although mostly dead colonies occur at the base of the island's hallow calm-water reefs, living colonies are never found on the uppermost surfaces of the reef flats. In contrast, M. annularis is common in comparable biotopes of continually submerged reefs at Jamaica (Goreau, 1959), Andros (Newell et al, 1959), Barbados (Lewis, 1960), and Isla de Lobos (Rigby and McIntire, 1966). Therefore, the absence of M. annularis from the occasionally emergent portions of Bonaire's reefs might be accepted as an indication of the inability of these corals to endure exposure. However, Milliman's (1969) is not the only report of emergence of M.
Click to view image in JPEG format. Fig. 5. [Grey Scale] Protruding branches of Acropora palmata, A. prolifera, and A. cervicornis at rear of A. palmata-dominated reef during low water in early afternoon, March 31, 1974. Normally, level of water is slightly above middle of wave- and/or organism-cut notch visible along low cliff. View to southeast from Karpata, Bonaire.
{kind=link}
End_Page 161------------------------
annularis, as it is the predominant coral species in the exposed Tortugas reef mentioned and illustrated by Vaughan (1919). Unfortunately, neither Milliman's nor, contrary to the belief of Glynn (1973a), Vaughan's studies included observations of the tolerance of the species to natural exposure. M. annularis was, however, not included by Vaughan (1913, 1919) among the corals that were experimentally found to be capable of surviving prolonged intervals out of water. On the other hand, unless it is assumed that the low-water periods during which Milliman and Vaughan conducted their investigations were unprecedented in their study areas, the simple presence of M. annularis makes obvious that the species has some degree of tolerance to being a wash, if only for infrequen and short intervals of time. Therefore, the prohibition of growth of M. annularis in the Diploria clivosa Zone (presumably by eradication of newly settled planulae, as not even exceedingly small coralla of dead M. annularis colonies have been found in this zone) and the decline of the species in the shallow calm-water reefs off Bonaire are probably attributable to extreme low-water periods that were repeated during the intense heat of midday over several consecutive days. These are the conditions that were previously suggested as responsible for truncation of Diploria strigosa colonies.
Although it seems reasonable to assume that D. strigosa polyps are somewhat better able to survive exposure than those of M. annularis, differences in the growth forms of the two corals must also greatly influence their relative endurance. Lewis (1960) has shown that the upward growth of the multilobed coralla of M. annularis causes the individual lobes to develop into elongated columns that are readily weakened by boring organisms, broken, and toppled. I have previously (Scatterday, 1974) attributed the destruction of M. annularis in shallow water off Bonaire to the susceptibility of the columnar growth form to bioerosion. Indeed, the various stages in the destruction of M. annularis described by Lewis can be observed affecting the tall co onies that grow in the deeper seaward parts of the channels between some of Bonaire's shallow calm-water reefs. However, many of the dead or mostly dead M. annularis colonies that now provide a foundation for the reefs grew in water too shallow to permit the growth of columns to unstable heights. Therefore, it seems probable that the destruction of the foundation colonies was initiated by their growth to the height at which polyps on the upper surfaces of centrally located lobes were killed during an interval of greater-than-usual exposure. Because the laterally directed growth of peripheral lobes would have caused them to be readily detached as bioerosion weakened the "core" of the coralla, all but a few lobes would ultimately be destroyed. Multer (1970, Figs. 5-20, 5-21, 5-22) has illustrated various stages in the destruction of small-lobed coralla of M. annularis similar to stages in the sequence proposed here. However, off Florida, where Multer's observations were made, the cause of death of the higher central portions of the colonies is most probably sediment accumulation rather than emergence. Whether the lobes are extended into columns or remain small, it seems clear that the multilobed growth form of M. annularis colonies enhances their susceptibility to destruction. In contrast, the nonlobate hemispherical form typical of Diploria strigosa colonies makes them far less susceptible to similar disintegration following death and truncation or even excavation of their tops, and therefore their survival as "microatolls" is common. >
Additional evidence supporting the probability that exposure has caused or at least initiated the destruction of M. annularis and inhibited its subsequent growth in extremely calm shallow water is provided by the nature of several reefs that lie in equally calm, but slightly deeper water. The tops of these reefs, which reach no closer to the surface than 1.5 m during extreme low water, are covered almost exclusively by an exceedingly profuse growth of healthy M. annularis. The most striking of these reefs (Fig. 6) is located in the wave-protected seaward part of a boca or estuary, Plaj'i Funchi, on Bonaire's northwest coast. The landward portion of the estuary has been sealed off from the sea for many years by a ridge of coral rubble, and a typical Diploria clivo a Zone assemblage between the rubble ridge and the reef attests to the absence of any unusual environmental conditions at this locality. As all other conditions appear
End_Page 162------------------------
to be identical to those under which the shallower reefs of diverse corals have developed, the slightly greater depth that allows the Plaj'i Funchi reef to escape emergence must account for the overwhemling predominance of living M. annularis in its construction. Moreover, the association of unusually high coral diversity with the shallow calm-water reefs that do experience emergence suggests that such diversity is related either directly to exposure or, more likely, to the decimation of M. annularis for which exposure is believed to have been responsible.
EFFECT OF REDUCTION OF MONTASTREA ANNULARIS
Two thirds (32 of 48) of the scleractinian species found to date off Bonaire are involved in the construction of the shallow calm-water reefs that are subject to emergence (Table 2). Such diversity (i.e., number of different species, relative abundance of each species has not been determined, but all are fairly common) is, as indicated previously, exceedingly unusual in shallow water in the Caribbean. It is usually found only in deeper water, especially at the depths occupied by the Montastrea annularis Zone (10 to 30 m off Bonaire). The reefs off the leeward coast of Barbados are perhaps
Click to view image in JPEG format. Fig. 6. [Grey Scale] Seaward portion of Montastrea annularis-dominated reef off Plaj'i Funchi on Bonaire's northwest coast. Such reefs are believed to be the result of the ability of M. annularis to exclude almost all other corals in settings where neither emergence nor steeply sloping substrates check its proliferation. Depth over tops of colonies at normal water level approximately 2 m. Density of M. annularis is greater nearer shore, but photographically overlooking the reef is prohibited by the shallow water in this location.
{kind=link}
Table 2. Corals on Shallow Calm-Water Reefs, Bonaire
End_Page 163------------------------
most similar to Bonaire's shallow calm-water reefs. However, Lewis (1960) listed only 17 species on the reef flats ("reef crest" in his terminology), including their flanks, and he recorded the prominence of living M. annularis. Moreover, as I have noted elsewhere (Scatterday, 1974), the assemblage of the shallow calm-water reefs off Bonaire resembles very closely the coral constituents of the "deep-water communities" off Barbados that Lewis described. Because of the shallow and wave-protected locales in which they customarily grow, the patch reefs found in various parts of the Caribbean or Gulf of Mexico also invite comparison with Bonaire's shallow calm-water reefs. However, these also differ from Bonaire's reefs in the occurrence of fewer species and in the prevalence of l ving M. annularis (e.g., Kornicker and Boyd, 1962; Milliman, 1973; Shinn, 1963).
Observations demonstrate that Montastrea annularis was the predominant living coral at or shortly after the inception of Bonaire's shallow calm-water reefs. Although coralla of species other than M. annularis are found at the base of the reefs, the amount of skeletal material contributed to the reef foundations by M. annularis greatly exceeds that provided by all other pioneering species and, for that matter, probably exceeds that which has been added recently by the extraordinarily heterogeneous corals that form the superjacent portions of the framework. Despite its prevalence at an early stage in the development of the reefs, M. annularis apparently could not attain the same degree of space monopolization as in the Plaj'i Funchi reef before enlarg ment of the colonies brought them near the surface at low water, thereby initiating their decline. It is probable that the decimation by exposure of the early colonies of M. annularis initially promoted diversity simply by providing a large expanse of suitable substrate on which a great variety of corals could settle without competition for space. Among these corals would have been some newly established colonies of M. annularis and, of course, those portions of the pioneering colonies that had escaped destruction would have continued their growth. Because M. annularis polyps may destroy by extracoelenteric digestion some other corals that are unfortunate enough to grow within reach of their extended mesenterial filiments (Lang, 1971, 1973) and may eliminate ma y species with which M. annularis colonies are competing for space, continued growth of colonies could have caused diversity to decline as the reef became more crowded. Had growth of M. annularis colonies not been periodically stopped or inhibited by emergence, diversity ultimately could have become no greater than that encountered in other continually submerged and M. annularis-dominated reefs in the Caribbean, or, under optimum conditions for proliferation, M. annularis could have monopolized almost all available space, attaining a climax in assemblage succession such as that found at Plaj'i Funchi.
The preceding explanation for the unusual diversity on some of Bonaire's reefs is comparable in certain respects to those advanced by investigators whose studies have involved comparisons of coral diversity on Indo-Pacific reefs. However, several assumptions that have been, or could be, made about Caribbean reefs and reef corals and that argue against the conclusions of this report should be considered.
If selective destruction of Montastrea annularis by emergence is indeed responsible for enhancement of diversity in shallow zones, it might be assumed that maximum diversity in deeper water should also be correlated with sparse occurrence of M. annularis. That this is not the case is made obvious by the name "Montastrea annularis Zone" for the deep-water assemblage in which the most varied corals are usually found. The occurrence of greatest diversity in this zone is due in part to the fact that a number of species (almost all of the one third of Bonaire's corals that are not found on the shallow calm-water reefs) are restricted to habitats with low levels of illumination, whereas most species that inhabit brightly lit, shallow-water habitats can ex st equally well at considerable depths. Of greater importance in determining diversity in the M. annularis Zone, however, is the fact that, unlike most of the shallow-water associates of M. annularis, many of the deeper-water corals, especially those belonging to the family Mussidae, have been found by Lang (1973) to be superior to M. annularis
End_Page 164------------------------
in the extracoelenteric-digestion hierarchy (i.e., M. annularis is eliminated by the mussids and corals of several other families in the same manner in which it disposes of its shallow-water subordinates; for details, see Lang, 1971, 1973). Occasional, local destruction of M. annularis also occurs in deeper water through toppling of bioeroded coralla, and the attrition brought about in this manner continually provides space that can be occupied, at least temporarily, by species that are lower in the extracoelenteric-digestion "pecking order." Indeed, Porter (1974) has assumed that the rate of such renewal of space is sufficient to prevent any one of the many contending corals from monopolizing space on Caribbean reefs. His assumption is based on the fact that the mor aggressive Caribbean corals (i.e., those able to destroy by digestion a large number of less aggressive corals) have a growth rate that is slow in relation to that of their subordinates. In contrast, the most aggressive Indo-Pacific corals (several of the representatives of Pocillipora) also have the fastest growth rate, a combination of traits that enables them to kill competitors by either digesting or overshadowing them, or both, and to quickly establish climax assemblages consisting only of congeners and characterized by dense growth (Porter, 1972a, c, 1974; Grigg and Maragos, 1974). In support of the belief that no Caribbean coral is sufficiently aggressive and fast growing to achieve the degree of space monopolization observed in the Pacific, Porter (1974) has shown that he highest diversity on Caribbean reefs off Panama occurs in areas of highest coral density where aggressive interactions should be at a maximum. Undoubtedly, the slow tendency toward space monopolization by M. annularis is offset by its rate of destruction by bioerosion-induced collapse along the abrupt edges of cliffs where the highest density and diversity occur on Panamanian reefs (Porter, 1972b) and on the steeply inclined slopes where the M. annularis Zone is found off Bonaire. For this reason, as well as the subordinate position of M. annularis in relation to a number of deep-water corals, high diversity can be maintained in the Montastrea annularis Zone. The rate of destruction by bioerosion in the M. annularis Zone is clearly faster, owever, than in the shallow water where Bonaire's M. annularis climax reefs are found.
The very gently sloping surface of the wave-and/or organism-eroded bench on which Bonaire's shallow reefs have formed has permitted greater reef development on the bench than on the steeply sloping flanks of the island (Scatterday, 1974). On steep slopes, toppling of bioerosion-weakened coralla is faster in part simply because of the more unstable attitudes that coralla must assume. In addition, the colonies and lobes of colonies of M. annularis in the shallow, but never emergent, reefs on the nearly level bench are so tightly appressed that, even if their bases are eroded through, they are supported upright by adjoining colonies or lobes. Only in a few places, such as along the margins of the reefs or where colonies have not completely coalesced, have portions of coralla bee even so much as tilted before coming to rest against neighboring coralla (Fig. 7). The density of living M. annularis on the shallow, continually submerged reefs is far greater than the density of all corals on either the occasionally emergent reefs or in the M. annularis Zone. Only scarce, small clusters of a few other scleractinians or of Millepora interrupt the otherwise continuous colonies that make up the M. annularis-dominated reefs, thereby further attesting to the negative influence of M. annularis on diversity where conditions have permitted colonies of the species to develop a climax assemblage.
DISCUSSION
In contrast to the situation observed off Bonaire, the seemingly adverse conditions that are associated with emergence usually have not been found to be conducive to coral diversity (Porter, 1972b). However, at least one other instance of emergence leading to results similar to those reported here has been observed (Fishelson, 1973). Moreover, although they do not deal with emergence, several other investigators (Grigg and Maragos, 1974; Porter, 1972a, c, 1974) have demonstrated that coral diversity may actually be increased by certain kinds of environmental adversity.
End_Page 165------------------------
On the basis of observations in the Red Sea, Fishelson (1973) has attributed unusually high diversity on some reefs to sporadic, unpredicted, extreme low tides that kill all or part of the corals, thereby preventing crowding and other unspecified forms of competition for space that might lead to decreased species heterogeneity. He has found that other reefs in the region that do not experience emergence possess considerably less diverse coral populations. Destruction or prevention of settlement of corals by "exposure," not to air but to high seas and accompanying abrasion by sediment, has been shown by Grigg and Maragos (1974) to be the cause of low density and high diversity of corals on submerged lava flows off Hawaii. The opposite relationship between density and diversity was foun on coeval flows in more hospitable, protected areas. These investigators also observed that, under otherwise similar environmental conditions, lower density and greater diversity occur in the pioneering assemblages on recently extruded flows than in the long-established climax assemblages of Pocillopora on the oldest flow.
The pernicious sporadic events or continuous conditions--subaerial exposure, exposure to damaging waves and sediment scour, and invasion by lava--to which Fishelson (1973) and Grigg and Maragos (1974) appeal in explaining coral diversity apparently do not selectively destroy only certain coral species. Hence, diversity is maintained simply because space adequate to ensure avoidance of encounters between superior and subordinate competitors is either periodically or constantly renewed. Of course, the consequences of exposure to corals off Bonaire is not nearly as catastrophic as that suffered during emergence of some Red Sea reefs or that resulting from incursions of lava. Creation of extensive, unoccupied space through widespread mortality of many different species does not result, an only Montastrea annularis appears to be severely affected.
Click to view image in JPEG format. Fig. 7. [Grey Scale] Close view of M. annularis on reef off Plaj'i Funchi. Although sector consisting of number of lobes has been detached from corallum in foreground, it has not fallen to a position in which the polyps that comprise the living "caps" of the lobes will be killed; rather, it is supported against neighboring colony in background. In central portions of reef, colonies have coalesced so completely that bioeroded lobes cannot be dislodged even to extent seen here. "Caps" range from 10 to 30 cm in diameter.
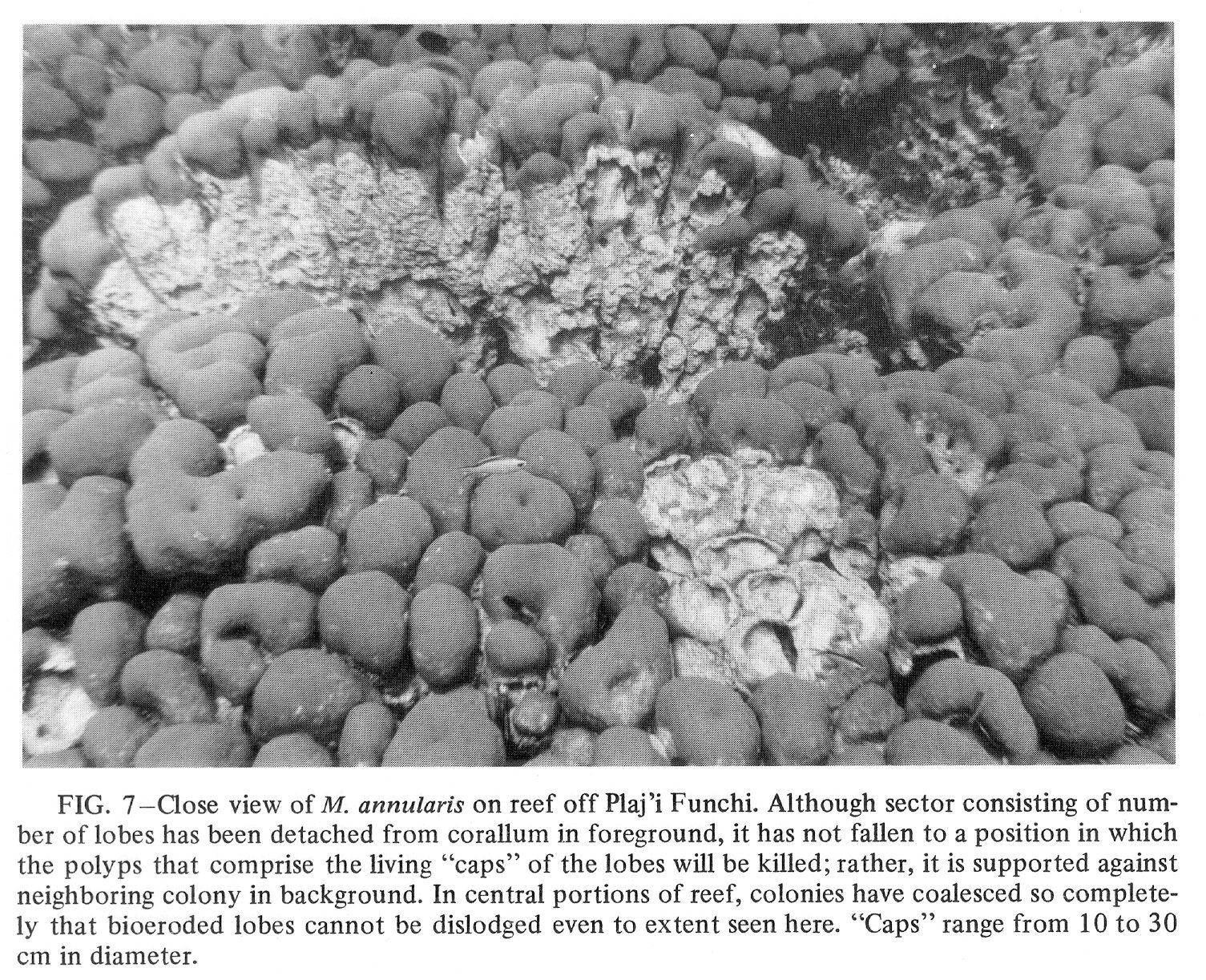{kind=link}
End_Page 166------------------------
Because it is also selective (although it is a biological agent instead of a physical event), the cause of enhanced diversity on some reefs off the Pacific coast of Panama is more comparable to the one that I have proposed for the unusual diversity on some of Bonaire's reefs. Porter (1972a, c, 1974) has shown that greater coral diversity occurs on Panamanian reefs infested by Acanthaster (the "crown-of-thorns" starfish) than on those from which the predator is absent. Acanthaster selectively feeds on Pocillopora damicornis, a coral that surpasses all others on the reefs in competition for space, not only by its rapid rate of growth, but also by its aggression in disposing of subordinate species by extracoelenteric digestion. Consequently, if unchecked by rapidly excludes all other corals and forms climax assemblages similar to those that the slower growing and more moderately aggressive M. annularis has formed at the level just below that reached by extreme low tides off Bonaire.
CONCLUSIONS
1. Low-water emergence of shallow-water reef biotopes that are inhabited by heterogeneous coral assemblages occurs in the Caribbean as well as in the Indo-Pacific. However, the occurrence of such emergence in the Caribbean is more sporadic and has been well documented at only three localities.
2. Although additional studies are needed to assess fully the effects of emergence on Caribbean corals, observations off Bonaire suggest that more than 20% of all known Caribbean scleractinians, as well as the hydrozoan Millepora, are able to withstand exposure for several hours in intense midday heat even without occasional external wetting by immersion beneath intermittent waves or by wave splash.
3. Montastrea annularis, a coral common in both shallow and deep water in the Caribbean,apparently is affected adversely by emergence. The inability of M. annularis to withstand subaerial exposure may be due in part to the relatively low tolerance of its polyps to heat and desiccation. However, the form of M. annularis coralla, which makes them especially susceptible to bioerosional disintegration following death of polyps on the more frequently exposed coralla summits, is believed to be the principal cause of the decline of representatives of the species after inordinately severe episodes of emergence.
4. Because M. annularis may exterminate by extracoelenteric digestion many of the corals with which it must compete for space in shallow water, the near elimination of these aggressive corals from emergence is believed to have resulted in the great diversity of corals that occurs on Bonaire's sometimes emergent reefs.
5. On shallow benches adjacent to the shore, where toppling of bioeroded coralla is prevented by dense growth and negligible substrate declivity and where water depth is just sufficient to permit development of reefs that escape protrusion above water during low-water episodes, M. annularis has been able to form reefs from which other corals have been almost entirely excluded.
6. In agreement with the results of other recent studies, observations off Bonaire have demonstrated that rather than eliminating all but one or two tolerant species, environmental rigors may actually enhance the diversity of a coral assemblage.
NOTE ADDED IN PROOF
Several of the conclusions of this report rest on the inference that the corals inhabiting shallow water off Bonaire are occasionally killed as a result of prolonged periods of subaerial exposure. Although I present evidence that justifies the inference, I call attention to the fact that none of the observed intervals of exposure were of sufficient duration to cause coral mortality. Subsequent to writing the report, however, I witnessed an episode of annihilation of the sort that had been inferred. Throughout a 15-day period in March 1976, Bonaire's shallow reef biotopes were awash during most of the daylight hours. As a result, all polyps of wholly emergent colonies and the polyps on upper portions of partially emergent colonies were killed. Exposed polyps showed signs of severe stre s after 9 days; none survived the next several days.
End_Page 167------------------------
References:
Bak, R. P. M., 1977, Coral reefs and their zonation in Netherlands Antilles: this volume.
Fishelson, L., 1973, Ecological and biological phenomena influencing coral-species composition on the reef tables at Eilat (Gulf of Aqaba, Red Sea): Marine Biology, v. 19, p. 183-196.
Glynn, P. W., 1968, Mass mortalities of echinoids and other reef flat organisms coincident with midday, low water exposures in Puerto Rico: Marine Biology, v. 1, p. 226-243.
Glynn, P. W., 1972, Observations on the ecology of the Caribbean and Pacific coasts of Panama: Biol. Soc. Washington Bull. 2, p. 13-30.
Glynn, P. W., 1973a, Aspects of the ecology of coral reefs in the western Atlantic region, in O. A. Jones and R. Endean, eds., Biology and geology of coral reefs, vol. 2; biology 1: New York, Academic Press, p. 271-324.
Glynn, P. W., 1973b, Ecology of a Caribbean coral reef--the Porites reef-flat biotope, part I, meteorology and hydrography: Marine Biology, v. 20, p. 293-318.
Goreau, T. F., 1959, The ecology of Jamaican coral reefs; I, species composition and zonation: Ecology, v. 40, p. 67-90.
Grigg, R. W., and J. E. Maragos, 1974, Recolonization of hermatypic corals on submerged lava flows in Hawaii: Ecology, v. 55, p. 387-395.
Kissling, D. L., 1965, Coral distribution on a shoal in Spanish Harbor, Florida Keys: Bull. Marine Sci., v. 15, p. 599-611.
Kornicker, L. S., and D. W. Boyd, 1962, Shallow-water geology and environments of Alacran reef complex, Campeche Bank, Mexico: AAPG Bull., v. 46, p. 640-673.
Lang, J., 1971, Interspecific aggression by scleractinian corals; 1, the rediscovery of Scolymia cubensis (Milne Edwards and Haime): Bull. Marine Sci., v. 21, p. 952-959.
Lang, J., 1973, Interspecific aggression by scleractinian corals; 2, why the race is not only to the swift: Bull. Marine Sci., v. 23, p. 260-279.
Lewis, J. B., 1960, The coral reefs and coral communities of Barbados, W. I.: Canadian Jour. Zoology, v. 38, p. 1133-1145.
Milliman, J. D., 1969, Four southwestern Caribbean atolls--Courtown Cays, Albuquerque Cays, Roncador Bank and Serrana Bank: Atoll Research Bull. 129, p. 1-22.
Milliman, J. D., 1973, Caribbean coral reefs, in O. A. Jones and R. Endean, eds., Biology and geology of coral reefs, vol. 1; geology 1: New York, Academic Press, p. 1-50.
Multer, H. G., 1970, Field guide to some carbonate rock environments: Miami Geol. Soc., 158 p.
Newell, N. D., and J. K. Rigby, 1957, Geologic studies on the Great Bahama Bank, in R. J. Le Blanc and J. G. Breeding, eds., Regional aspects of carbonate deposition: SEPM Spec. Pub. 5, p. 15-79.
Newell, N. D., et al, 1951, Shoal-water geology and environments, eastern Andros Island, Bahamas: Am. Mus. Nat. History Bull., v. 97, art. 1, p. 5-29.
Newell, N. D., et al, 1959, Organism communities and bottom facies, Great Bahama Bank: Am. Mus. Nat. History Bull., v. 117, art. 4, p. 181-228.
Porter, J. W., 1972a, Ecology and species diversity of coral reefs on opposite sides of the Isthmus of Panama: Biol. Soc. Washington Bull. 2, p. 89-116.
Porter, J. W., 1972b, Patterns of species diversity in Caribbean reef corals: Ecology, v. 53, p. 745-748.
Porter, J. W., 1972c, Predation by Acanthaster and its effect on coral species diversity: Am. Naturalist, v. 106, p. 487-492.
Porter, J. W., 1974, Community structure of coral reefs on opposite sides of the Isthmus of Panama: Science, v. 186, p. 543-545.
Rigby, J. K., and W. G. McIntire, 1966, The Isla de Lobos and associated reefs, Veracruz, Mexico: Brigham Young Univ. Geol. Studies, v. 13, p. 3-46.
Roos, P. J., 1964, The distribution of reef corals in Curacao: Studies Fauna Curacao, v. 20, p. 1-51.
Scatterday, J. W., 1974, Reefs and associated coral assemblages off Bonaire, Netherlands Antilles, and their bearing on Pleistocene and Recent reef models: 2d Internat. Coral Reef Symposium Proc., v. 2, p. 85-106.
Shinn, E., 1963, Spur and groove formation on the Florida reef tract: Jour. Sed. Petrology, v. 33, p. 291-303.
Storr, J. F., 1964, Ecology and oceanography of the coral-reef tract, Abaco Island, Bahamas: Geol. Soc. America Spec. Paper 78, 98 p.
Vaughan, T. W., 1913, Studies of the geology and of the madreporaria of the Bahamas and of southern Florida: Carnegie Inst. Washington Yearbook 11, p. 153-162.
Vaughan, T. W., 1919, Corals and the formation of coral reefs: Smithsonian Inst. Ann. Rept. for 1917, p. 189-238.
End_Page 168------------------------
Wells, J. W., 1957, Coral reefs, in J. W. Hedgpeth, ed., Treatise on marine ecology and paleoecology; vol. 1, ecology: Geol. Soc. America Mem. 67, p. 609-631.
Wells, J. W., 1973, Two new hermatypic scleractinian corals from the West Indies: Bull. Marine Sci., v. 23, p. 925-932.
Wells, J. W., and J. C. Lang, 1973, Systematic list of Jamaican shallow-water scleractinian: Bull. Marine Sci., v. 23, p. 55-58.
Yonge, C. M., 1935, Studies on the biology of Tortugas corals; 2, variation in the genus Siderastrea: Carnegie Inst. Washington Pub. 452, p. 199-208.
Zaneveld, J. S., 1957, Micro-atolls in the Netherlands Antilles (abs.): Inter-Island Marine Biology Conf. Rept., Inst. Marine Biology, Univ. Puerto Rico, p. 18-19.
End_of_Record - Last_Page 169-------
Acknowledgments:
(2) Department of Geological Sciences, State University of New York, College of Arts and Science at Geneseo, Geneseo, New York 14454.
A grant-in-aid from the SUNY/Research Foundation helped defray field expenses in 1974.
Pay-Per-View Purchase Options
The article is available through a document delivery service. Explain these Purchase Options.
| Watermarked PDF Document: $14 | |
| Open PDF Document: $24 |